


Science Bulletin | 万向元团队发现ZmMs1反馈抑制环调控ZmACSN—PKSB—TKPR1复合体从而精确控制玉米花粉发育
花粉外壁(Pollen exine)是由孢粉素(Sporopollenin)组成的一种惰性的脂质层,位于花粉壁的最外层(Ariizumi et al., 2011; Shi et al., 2015),在花粉传播过程中对精子具有保护作用。作为花粉与柱头之间相互作用的关键界面,花粉外壁还决定了花粉的成功受精能力,进一步对作物产量产生直接影响,因此引起了植物科学界的广泛关注。该团队已发现ZmMs1介导的反馈抑制环通过调控6个孢粉素和3个活性氧(ROS)相关基因来确保花粉外壁形成(Hou et al., 2023)。然而,除了这9个靶基因,许多其他基因也可能是该反馈抑制环的靶标。因此,发掘更多ZmMs1反馈抑制环的靶基因可进一步加深对调控花粉外壁形成网络的理解。
2025年4月10日,北京科技大学生物农业研究院、北京中智生物农业国际研究院万向元团队在Science Bulletin(即时IF: 20.6)在线发表题为ZmMS1-orchestrated feedback repression loop regulates ZmACSN2/3—PKSB—TKPR1-1/-2 complex to ensure maize pollen development的研究论文。该研究通过CRISPR/Cas9编辑技术鉴定到两个参与孢粉素生物合成相关的GMS基因ZmACSN2和ZmACSN3,它们编码脂肪酰基辅酶A合成酶,具有对长链脂肪酸和长链羟基脂肪酸的催化活性。此外,ZmACSN2和ZmACSN3可以与ZmPKSB和ZmTKPR1-1/2形成复合物级联催化脂质生物合成。该代谢产物的形成和终止由ZmMS1介导的反馈抑制环调控,从而精确控制玉米花粉外壁的形成。
该团队基于CRISPR/Cas9介导的基因编辑技术,发现编码脂酰基CoA合成酶基因ZmACSN2和ZmACSN3单突完全可育,双突变体(acsn2; acsn3)表现为完全雄性不育,且花粉外壁发育缺陷(图1a,b),因此,ZmACSN2和ZmACSN3在雄花发育方面存在功能冗余。该团队前期克隆的ZmPKSB、ZmTKPR1-1/-2基因(Liu et al., 2022;An et al., 2023)与ZmACSN2/ZmACSN3在花药发育过程中呈现相似的表达模式,其编码蛋白同样定位于内质网,且pksb和tkpr1-1; tkpr1-2突变体花粉外壁也明显变薄。基于酵母双杂交、BiFC和Co-IP等多种方法证明:上述5个蛋白彼此互作形成多酶复合物(图1c)。已有报道表明多酶复合物通常在代谢途径中催化连续反应(Obata et al., 2019)。该研究进一步发现ZmACSN2/3、ZmPKSB和ZmTKPR1-1/-2能够以C16、C18、C16-OH为底物,进行高效的级联酶促反应以生成还原型四酮α-吡喃酮(图1d)。由此,新发现存在于玉米花药中调控孢粉素生物合成的蛋白复合物,ZmACSN2/3-ZmPKSB-ZmTKPR1-1/-2,可进行级联酶促反应以催化脂肪酸和脂肪醇最终形成孢粉素合成必要前体四酮α-吡喃酮(图1e)。
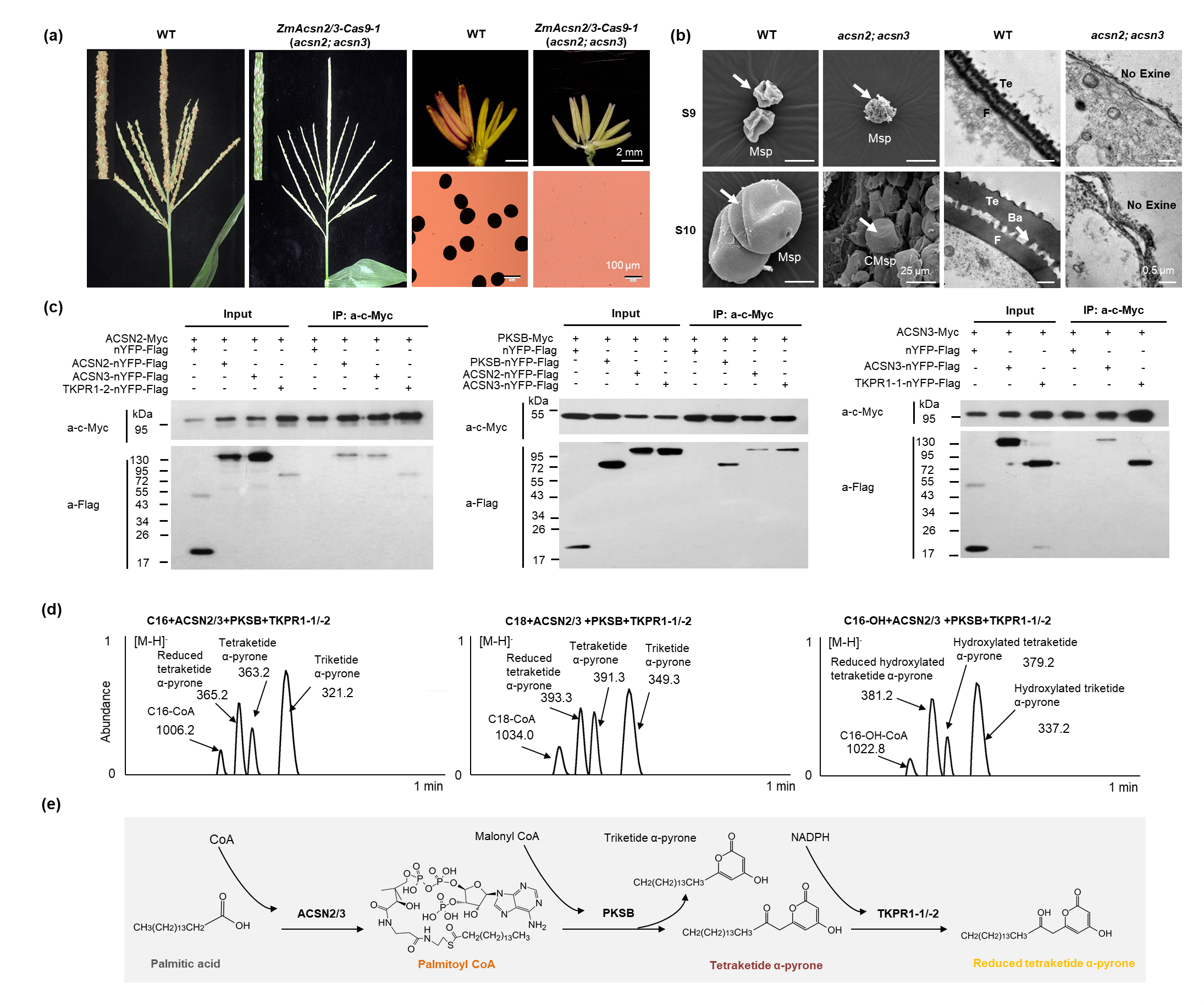
图1. ZmACSN2和ZmACSN3双突变体表型观察及ZmACSN2/3-ZmPKSB-ZmTKPR1-1/-2复合物酶活分析
该团队之前报道的ZmMS1介导的反馈抑制环(ZmbHLH51-ZmMYB84-ZmMS7-ZmMS1)精确调控花粉外壁形成(Hou et al., 2023)。该研究中发现ZmbHLH51, ZmMyb84, ZmMs7, 和ZmMs1在花药发育的S9时期与ZmAcsn2/3, ZmPksb, and ZmTkpr1-1/-2有共同的表达峰。qRT-PCR、TDLR、EMSA和ChIP-seq等实验证实ZmbHLH51、ZmMS7和ZmMS1可直接调控ZmAcsn2和ZmAcsn3表达,而ZmMYB84可直接激活ZmAcsn2/3、ZmPksb和ZmTkpr1-1/-2基因表达(图2a-j)。因此,ZmACSN2/3-ZmPKSB-ZmTKPR1-1/-2受ZmMS1介导的反馈抑制环精确调控(图2k)。总之,ZmMS1反馈抑制环可精确调控ZmACSN2/3- ZmPKSB-ZmTKPR1-1/-2蛋白复合物,促进孢粉素前体的及时形成和终止,最终形成转录调控通路和蛋白复合物之间的遗传网络,以确保花粉外壁发育。
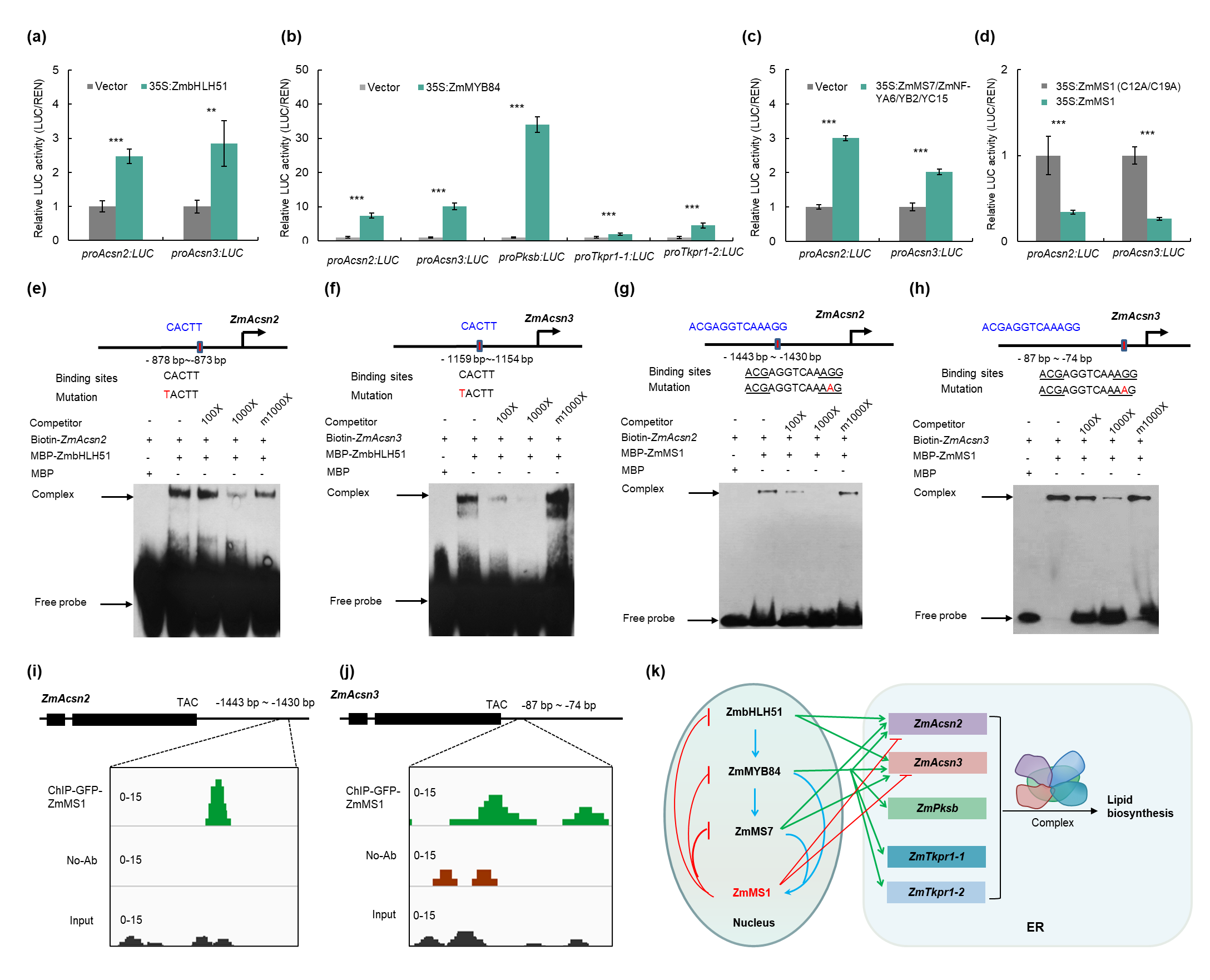
图2. ZmMS1反馈抑制环精确调控ZmACSN2/3-ZmPKSB-ZmTKPR1-1/-2蛋白复合物
该研究还发现,尽管ZmACSN2/3-ZmPKSB-ZmTKPR1-1/-2蛋白复合物可催化孢粉素生物合成的前体,但它们突变后对花粉外壁和花药角质层造成的影响却不完全一致,如acsn2; acsn3不能形成花粉外壁,tkpr1-1; tkpr1-2花粉外壁明显薄于pksb突变体;acsn2; acsn3可以形成很薄的花药外壁,tkpr1-1; tkpr1-2花药外壁光滑,然而pksb花药角质层则更加致密(图3a)。进一步分析发现,在acsn2; acsn3、pksb和tkpr1-1/-2突变体花药中,孢粉素、角质和蜡质相关基因的表达量以及角质、蜡质、内脂总含量和主要单体含量等也呈现明显差异(图3b),表明了植物花粉外壁形成过程中脂代谢途径的复杂性。据此,提出了转录因子ZmMS1反馈抑制环调控的ZmACSN2/3-ZmPKSB-ZmTKPR1-1/-2蛋白复合调控玉米花药和花粉发育的工作模型(图4),为研究植物脂类基因参与孢粉素、角质和蜡质合成与运输提供了新思路。
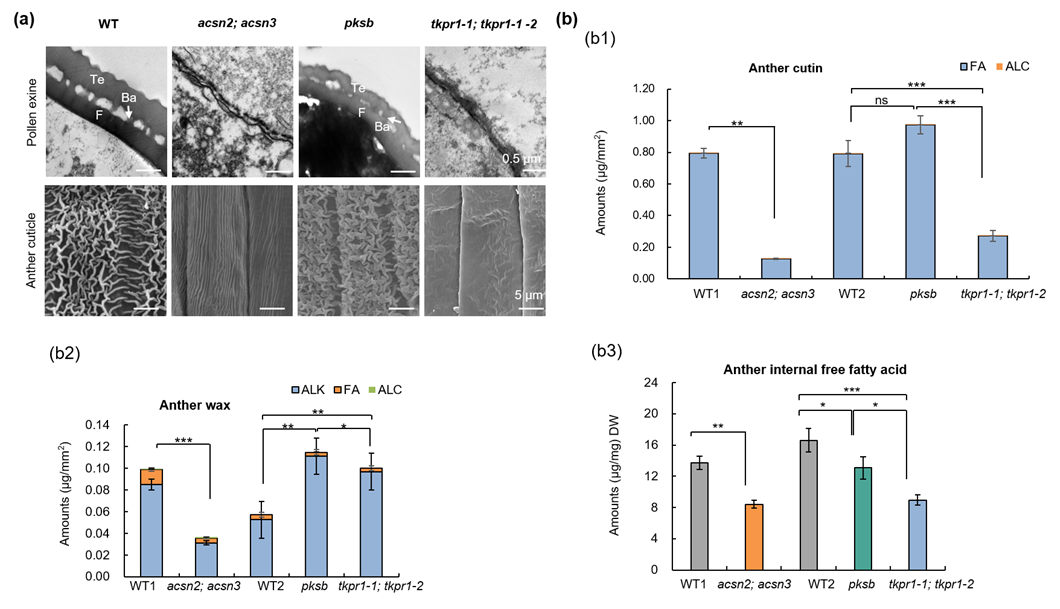
图3. ZmACSN2/3-ZmPKSB-ZmTKPR1-1/-2蛋白复合物中不同蛋白功能差异
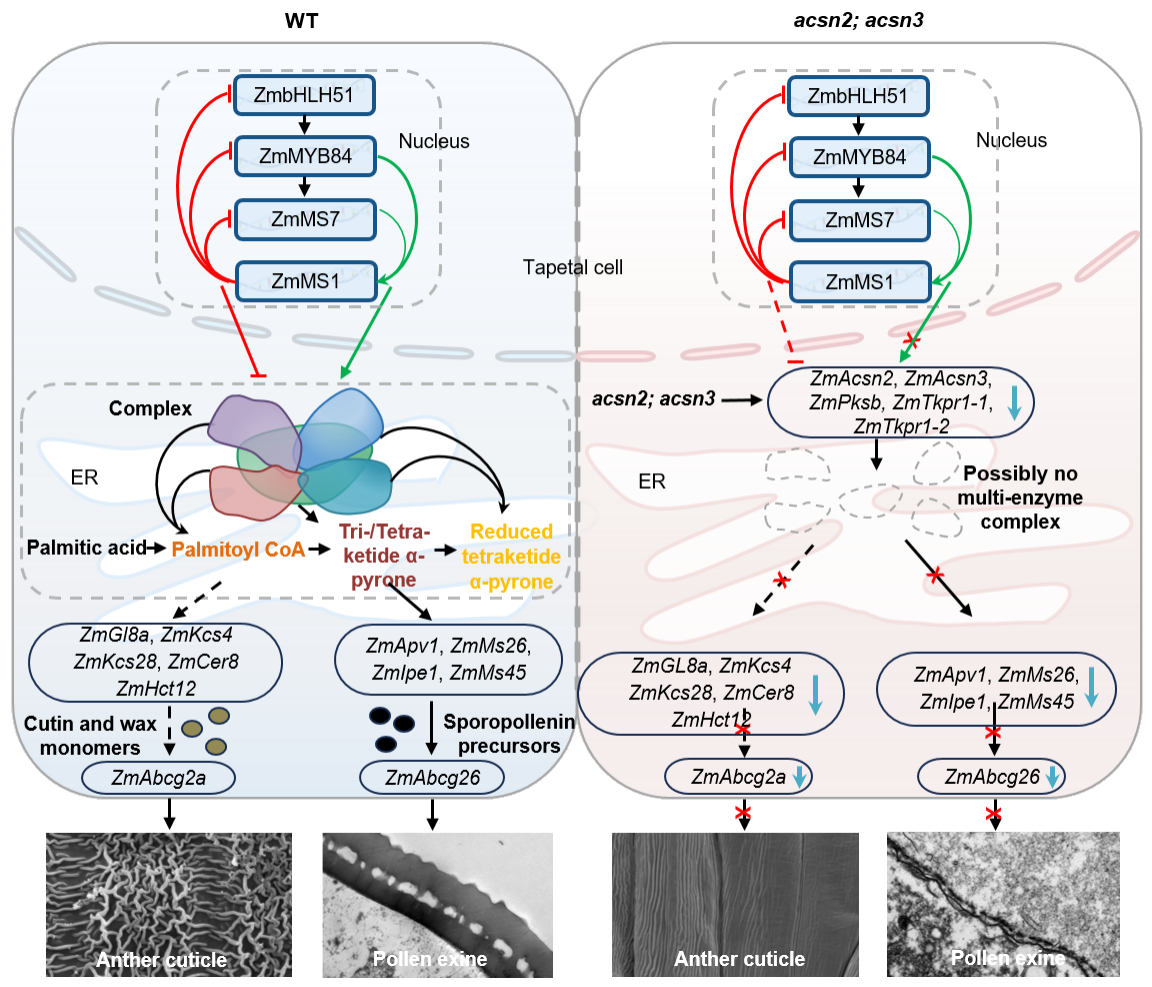
图4. ZmMS1反馈抑制环精确调控ZmACSN2/3-ZmPKSB-ZmTKPR1-1/-2蛋白复合物控制玉米花药和花粉发育的工作模型
北京科技大学生物农业研究院安学丽教授、已毕业博士生房超伟、张少伟为该论文共同第一作者,万向元教授为通讯作者。该研究得到了“十四五”国家重点研发计划、国家自然科学基金、中央高校基本科研业务费等项目的资助。
通讯作者简介:万向元,北京科技大学生物农业研究院院长、二级教授、博导,教育部长江学者特岗教授,国家“万人计划”领军人才,北京市特聘专家,“十四五”国家重点研发专项项目首席科学家。主要从事玉米雄性不育、绿色高效生物育种及其遗传基础研究;已在PNAS、Mol. Plant、Sci Bull.、Nat. Com.、Cell Res.、PBJ、JAR、JCLP等国内外期刊发表论文150余篇;参与出版教材和专著8部;授权国家发明专利42项;获批国家植物新品种权5项;以第1完成人获北京市技术发明奖一等奖、中国专利银奖、中国科协“科创中国”先导技术榜、大北农科技奖一等奖等科研/论文奖励13项。兼任北京中智生物农业国际研究院院长、中国作物学会理事、中国生物工程学会常务理事、全国作物杂种优势与生物育种学术大会执行主席、《Crop Journal》副主编等。
参考文献:
An X, Zhang S, Jiang Y, et al. CRISPR/Cas9-based genome editing of 14 lipid metabolic genes reveals a sporopollenin metabolon ZmPKSB-ZmTKPR1-1/-2 required for pollen exine formation in maize. Plant Biotechnol J 2024; 22: 216-232.
Ariizumi T, Toriyama K. Genetic regulation of sporopollenin synthesis and pollen exine development. Annu Rev Plant Biol 2011; 62: 437-460.191
Hou Q, An X, Ma B, et al. ZmMS1/ZmLBD30-orchestrated transcriptional regulatory networks precisely control pollen exine development. Mol Plant 2023; 16: 1321-1338.
Liu X, Jiang Y, Wu S, et al. The ZmMYB84‐ZmPKSB regulatory module controls male fertility through modulating anther cuticle—pollen exine trade‐off in maize anthers. Plant Biotechnol J 2022; 20: 2342-2356.
Obata T. Metabolons in plant primary and secondary metabolism. Phytochem Rev 2019; 18: 1483-1507.
Shi J, Cui M, Yang L, et al. Genetic and biochemical mechanisms of pollen wall development. Trends Plant Sci 2015; 20: 741-753.
原文链接:https://doi.org/10.1016/j.scib.2025.03.059