


Journal of Advanced Research | 发现育性新基因ZmMs13三峰值表达分别调控玉米雄花发育的三个不同生物学过程
植物花药和花粉发育是一个由大量基因协同控制的复杂生物学过程,相关基因的功能缺失往往导致细胞核雄性不育(Genic male sterility,GMS)。花药壁由四层体细胞组成,由外向内依次是表皮层、内皮层、中间层和绒毡层。其中,绒毡层对花粉发育至关重要,包括分泌胼胝质降解的酶,参与孢粉素、花药角质和蜡质前体的合成和转运,最终通过细胞程序性死亡(Programmed cell death,PCD)而降解,为花粉外壁的形成提供必需的营养物质。脂类物质在绒毡层合成后,由Lipid transfer protein(LTP)和ATP Binding Cassette G(ABCG)等脂转运蛋白运出绒毡层细胞,分别向外转运至花药外表皮合成角质层(cuticle),向内转运至小孢子表面形成花粉外壁(exine),二者对于雄性育性和花粉活力起重要的保护作用。前期研究表明ABCG转运蛋白参与花药角质层的各种蜡质和角质单体、花粉壁的孢粉素前体和含油层成分的转运[1];玉米基因组中共有54个ABCG基因[2],然而参与玉米花粉发育的ABCG基因较少报道[3],ABCG转运蛋白在植物花药和花粉发育中的精细调控机制尚不清楚。
2022年9月18日,北京科技大学、北京中智生物农业国际研究院万向元教授团队在国际著名综合性学术期刊Journal of Advanced Research(IF:12.822)在线发表了题为 “Triphasic Regulation of ZmMs13 Encoding an ABCG Transporter Is Sequentially Required for Callose Dissolution, Pollen Exine and Anther Cuticle Formation in Maize”的研究论文。首次发现玉米雄性不育新基因ZmMs13/ZmABCG2a在花药/花粉发育的前期(S5)、中期(S8b)和后期(S10)具有三个表达峰,分别受转录因子ZmbHLH122、ZmMYB84和ZmMYB33-1/-2调控,依次参与玉米花药/花粉的胼胝质降解、绒毡层PCD和花粉外壁发育、花药角质层形成等过程;而且ZmMS13蛋白的一个单氨基酸(I311)对其ATP酶活性至关重要,缺失导致完全雄性不育;这为解析ABCG转运蛋白在植物花药和花粉发育过程中的精细调控机制提供了新思路和新视角。

该团队利用2个玉米隐性核雄性不育突变体ms13和ms13-6060,通过图位克隆、DNA测序及基因编辑等方法鉴定了Zm00001d013960是育性控制基因ZmMs13,在ms13和ms13-6060中ZmMs13的第四外显子分别被插入20bp和缺失3bp,分别导致一个移码突变和单氨基酸缺失(I311)(图1)。ZmMs13基因在花药中特异表达,并且在花药发育的S5,S8b和S10时期各有一个表达峰;该基因编码一个内质网和质膜双定位的ABCG转运蛋白(ZmABCG2a),具有ATP酶和脂质结合活性,ms13-6060突变体中ZmMS13蛋白的单氨基酸缺失(I311)导致ATP酶活性急剧降低,表明I311是ATP酶活性的关键位点(图2)。
前期,该团队已经报道了18个转录因子(TF)GMS新基因在玉米花药不同发育时期的表达模式,其中有7个在S5时期有表达峰,2个在S8b时期有表达峰,2个在S10时期有表达峰[4]。本研究通过qPCR、Pro-LUC、EMSA等分析发现,ZmMs13/ZmABCG2a在花药/花粉发育的S5、S8b和S10三个时期具有三个表达峰,分别受转录因子ZmbHLH122、ZmMYB84和ZmMYB33-1/-2调控,对应3个生物学事件,具体详述如下:
第一峰值期:ZmbHLH122-ZmMs13调控模块参与花药发育早期的胼胝壁降解。为了验证ZmbHLH122对ZmMs13调控作用,通过qPCR,TDLR和EMSA实验证明ZmbHLH122在S5时期直接激活ZmMs13表达;胼胝质苯胺蓝染色结果显示,在ms13-6060突变体中,S7和S8a时期花药的胼胝质降解明显受阻(图3A);胼胝质降解相关基因ZmA6和ZmCDM1在突变体花药S5和S6时期表达显著下调,合成相关基因ZmCalS5在S6和S7时期表达显著上调,表明ZmMs13通过调控花药胼胝质相关基因的表达来影响胼胝质代谢。因此,在玉米花药发育早期(S5-S7),ZmbHLH122直接激活ZmMs13表达,进而调控胼胝质的降解。
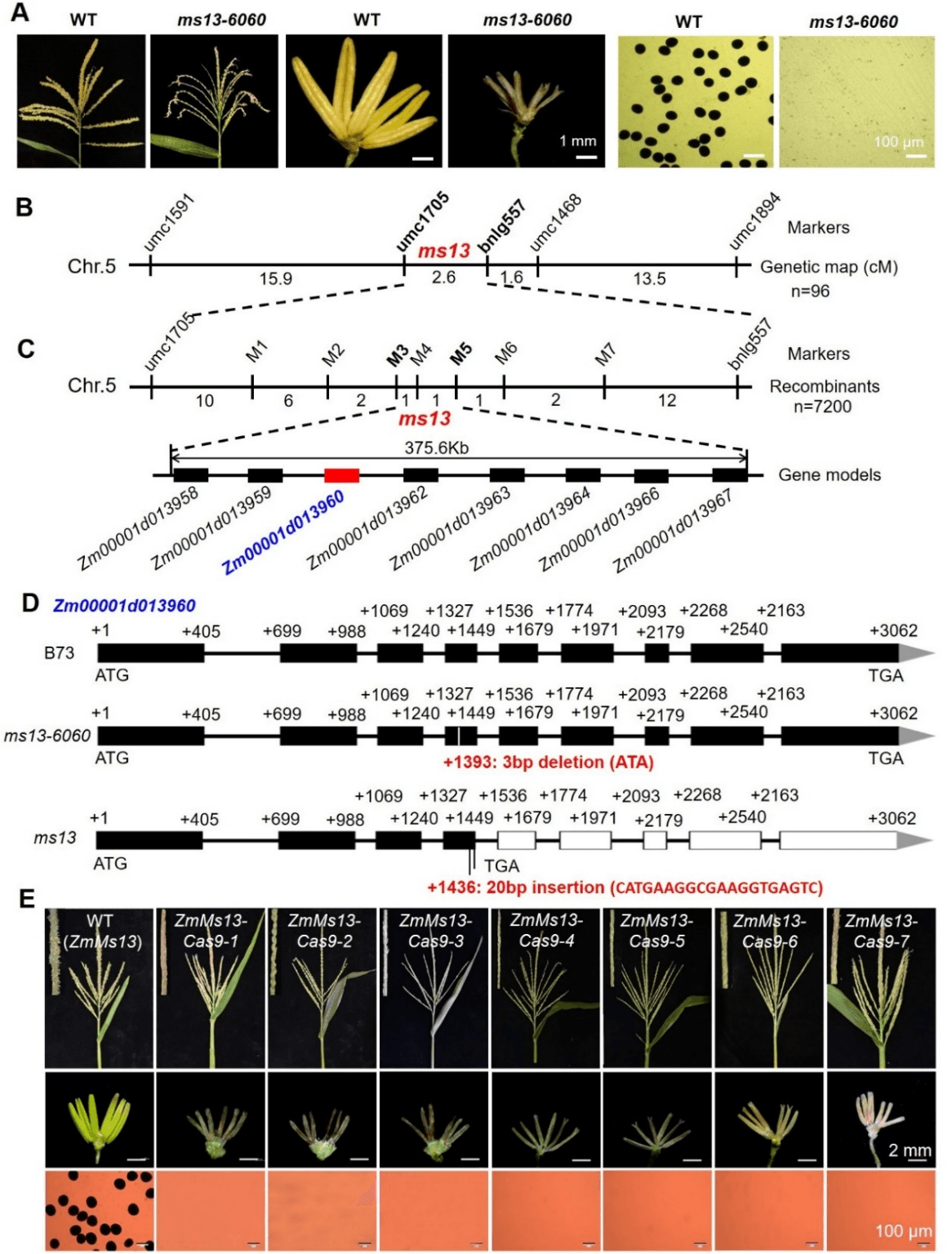
图1. 野生型和ms13-6060突变体表型、ZmMs13基因精细定位与图位克隆及基因编辑验证
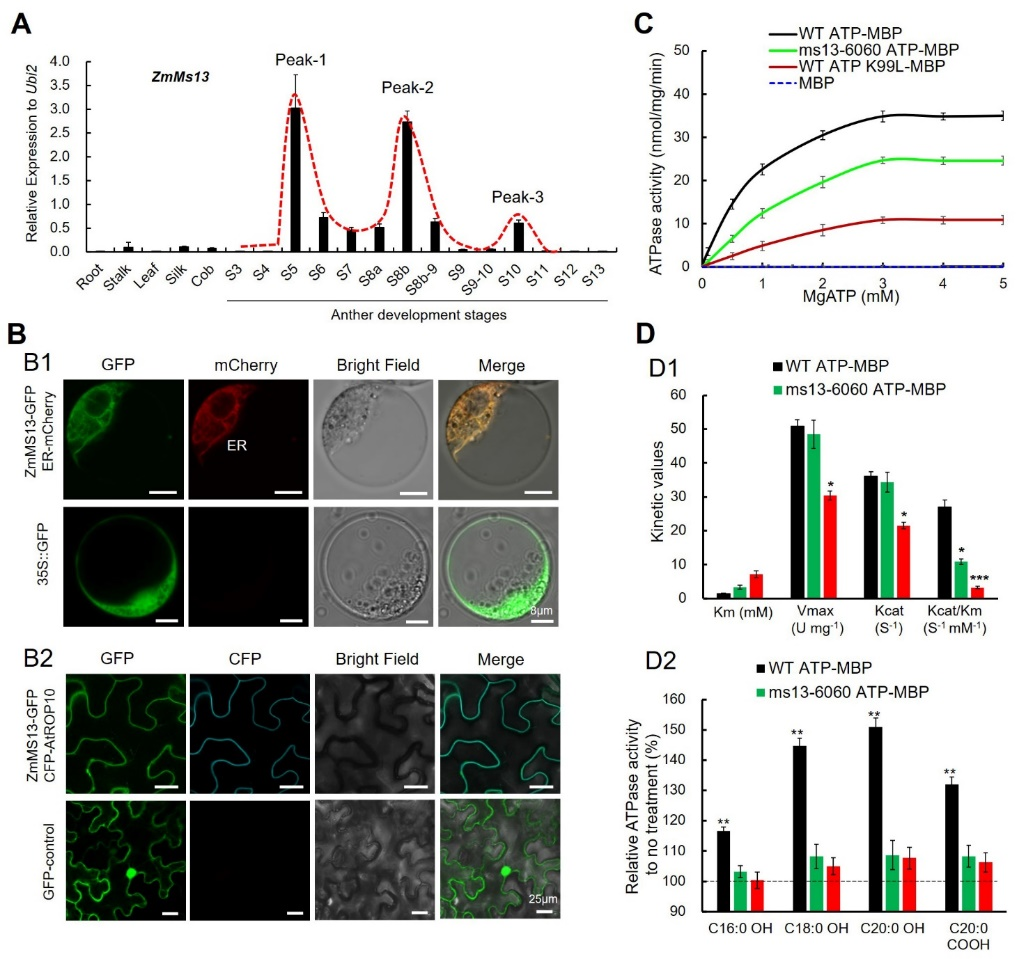
图2. ZmMs13基因时空特异表达、蛋白亚细胞定位与酶活分析
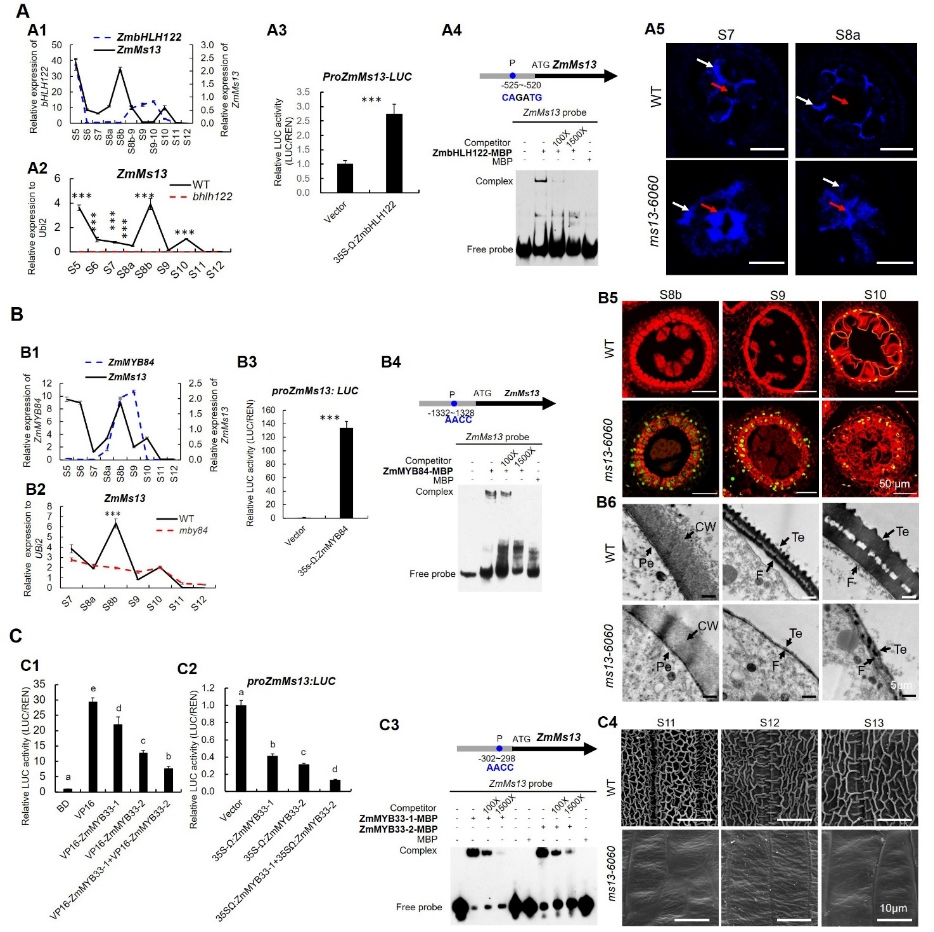
图3. ZmbHLH122、ZmMYB84和ZmMYB33-1/-2调控ZmMs13基因表达,进而影响玉米花药和花粉发育
第二峰值期:ZmMYB84-ZmMs13调控模块参与花药绒毡层PCD和花粉外壁形成。qPCR和Pro-LUC分析表明ZmMYB84与ZmMs13在S8b时期共表达,且ZmMYB84与ZmMs13的启动子直接结合并激活其表达;TUNEL 实验结果表明ms13-6060突变体绒毡层PCD提前发生(图3B);ROS清除相关基因ZmTRXb和ZmPODa在S7-S8a时期表达降低,但PCD相关基因ZmCP1表达升高;TEM分析显示突变体S9和S10期小孢子外壁比野生型(WT)异常变薄(图3B)。因此,ZmMs13在四分体时期(S8b)直接被ZmMYB84激活,并参与了ROS诱导的绒毡层PCD和花粉外壁形成过程。
第三峰值期:ZmMYB33-1/2~ZmMs13调控模块参与花药角质层形成。qPCR分析表明ZmMYB33-1/-2与ZmMs13在S10时期有共同的表达峰值;TDLR和EMSA分析表明,ZmMYB33-1和ZmMYB33-2直接协同抑制ZmMs13的表达;SEM和脂质染色结果表明,ZmMs13基因突变抑制花药角质层形成(图3C)。因此,ZmMYB33-1/-2在花药发育S10时期调控ZmMs13转录,影响玉米花药角质层形成。
总之,本研究发现玉米ZmMs13基因在花粉发育早期(S5),四分体期(S8b)和单核小孢子后期(S10),依次受转录因子ZmbHLH122、ZmMYB84和ZmMYB33-1/-2调控,并通过影响下游相关基因的表达,参与胼胝质降解,绒毡层PCD和花粉外壁以及花药角质层的形成,最终调控玉米雄花育性发育(图4)。该研究首次发现一个基因多峰值表达分别受不同转录因子调控,分别对应不同的生物学功能,且ZmMS13蛋白的一个单氨基酸(I311)对其ATP酶活性至关重要,缺失导致完全雄性不育;这将为植物ABCG转运蛋白和多峰值表达基因的分子机理解析提供参考思路。
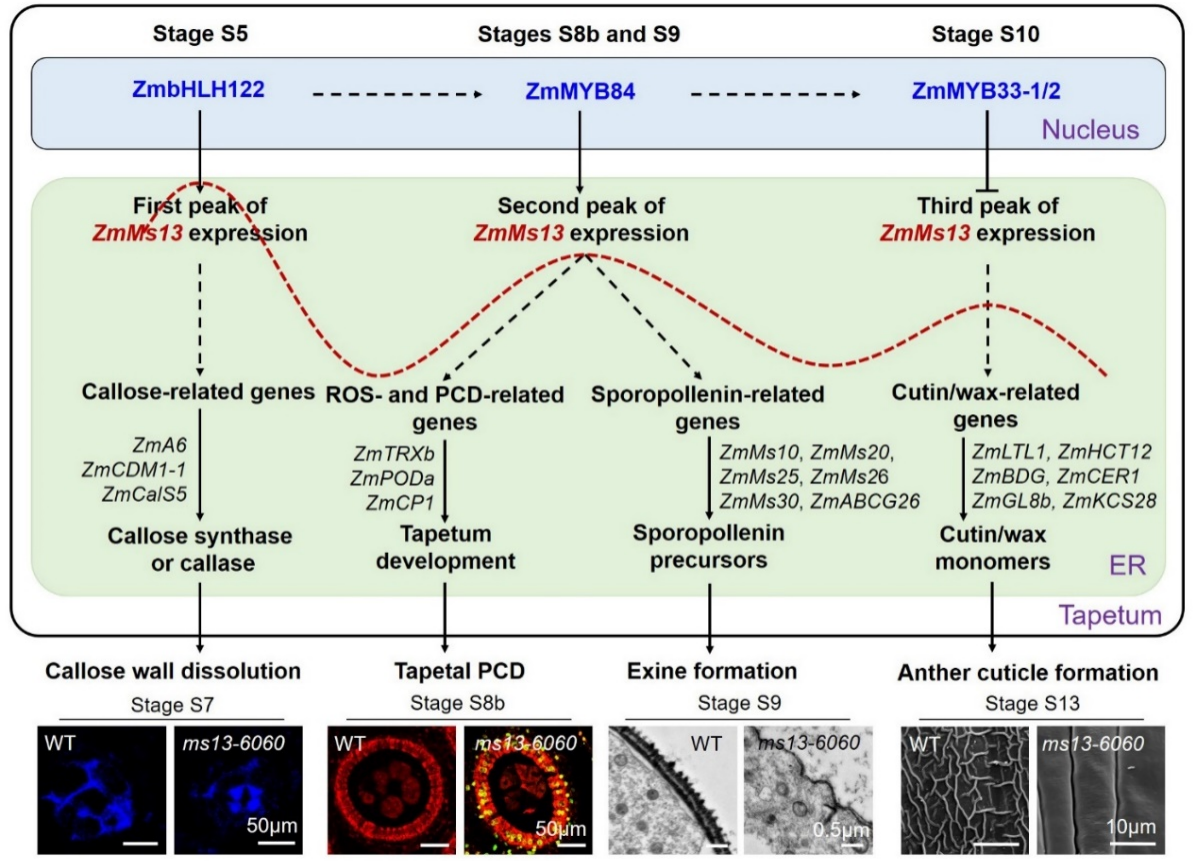
图4. 玉米ZmMs13分子调控机制及其影响玉米花药和花粉发育的工作模型
本研究主要依托北京科技大学生物与农业研究中心、北京中智生物农业国际研究院、北京首佳利华科技有限公司等单位完成,北京科技大学在读博士生房超伟、吴锁伟教授级高工和牛灿芳副教授为该论文共同第一作者,万向元教授为通讯作者。该研究得到了国家重点研发计划、国家自然科学基金、中央高校基本科研业务费等项目的联合资助。
原文链接:https://doi.org/10.1016/j.jare.2022.09.006
参考文献:
[1] Hwang JU, Song WY, Hong D, Ko D, Yamaoka Y, Jang S, et al. Plant ABC Transporters Enable Many Unique Aspects of a Terrestrial Plant's Lifestyle. Mol Plant 2016;9(3): 338-55.
[2] Wu S, Fang C, Li Z, Wang Y, et al. ATP-Binding Cassette G Transporters and Their Multiple Roles Especially for Male Fertility in Arabidopsis, Rice and Maize. Int J Mol Sci 2022; 23(16):9304.
[3] Jiang Y, Li Z, Liu X, Zhu T, Xie K, Hou Q, et al. ZmFAR1 and ZmABCG26 Regulated by microRNA Are Essential for Lipid Metabolism in Maize Anther. Int J Mol Sci 2021; 22(15): 7916.
[4] Jiang Y, An X, Li Z, Yan T, Zhu T, Xie K, et al. CRISPR/Cas9-based discovery of maize transcription factors regulating male sterility and their functional conservation in plants. Plant Biotechnol J 2021; 19(9): 1769-84