


NPR | 万向元团队与美国爱荷华州立大学Peters教授联合系统分析了植物萜类(二萜)进化
二萜类化合物属于天然化合物中种类丰富的一大类群,目前已经鉴定出的二萜天然产物至少12000种, 其中在植物中包括光合色素、植物激素、次生代谢产物等。二萜化合物结构的多样性源于简单的异戊二烯单元(C5)、异戊烯基二磷酸(isopentenyl pyrophosphate, IPP)和其异构体二甲丙烯二磷酸(dimethylallyl pyrophosphate, DMAPP)。目前,已有很多研究报道了二萜天然产物结构的多样性且具有不同的生理功能,在农业、林业、工业及医药领域均有重要的应用价值。
2022年12月06日,北京科技大学、北京中智生物农业国际研究院万向元教授、张娟副教授与美国爱荷华州立大学Peters教授团队,受邀联合在国际知名期刊Natural Product Reports(IF: 15.111)在线发表题为“Plant (di)terpenoid evolution: from pigments to hormones and beyond”的论文,系统分析了二萜天然产物合成前体、萜类合酶(TPS)家族、P450家族、下游的修饰酶以及合成基因簇,对深度理解二萜天然产物的合成及多样性具有重要意义。

首先,该论文介绍了二萜天然产物的顺式和反式合成前体:全反式(E,E,E,)-geranylgeranyl diphosphate(二磷酸香叶酰,GGPP)和全顺式(Z,Z,Z)-nerylneryl diphosphate (二磷酸内酰基,NNPP)是二萜合成的两个主要前体。其中NNPP是由cis-prenyl transferases (顺式戊烯基转移酶,CPTs)合成,而GGPP是由diphosphate synthases (二磷酸合酶,IDSs)合成。值得注意,通过对植物CPTs和IDSs家族分析,发现这些合成酶除了进行经典的“从头到尾”合成反应外,有些成员还通过缩合反应合成不规则的萜类(包括环丙基反应、环丁基反应等)(图1)。
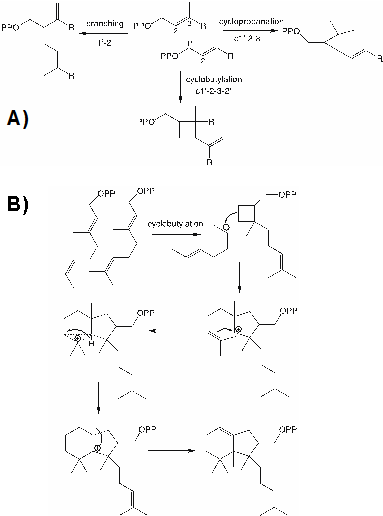
图1. 不规则的二萜合成
其次,对萜类合酶、P450家族及下游的修饰酶分别进行了系统分析。萜类合酶TPSs根据其功能域可分为两大类: Class I和II类。Class II类二萜合酶功能域是DXDD酸性活性中心和H-N碱性活性中心,这两个功能域在二萜合酶中均是保守的。GGPP在Class II类二萜合酶催化下, 选择性地生成带有特殊立体构象的环状烃类结构, 如copalyl diphosphate (二磷酸共苯酯, CPP), 目前已知的有nor-CPP、ent-CPP、syn-CPP和ent-syn-CPP四种不同的构象。GGPP初始质子化后, 在异戊烯基碳链上接着级联反应, 发生多步环化,最终被H-N结合的水分子组成的碱性活性中心终止,完成反应。位于中间位置的天冬氨酸是质子化反应的关键氨基酸,而H或N突变后均可导致不一样的产物(图2)。

图2. CPSs催化反应及关键氨基酸
Class I类环化酶包含非常保守的DDXXD和不是很保守的NSE/DTE功能域, 其催化反应涉及二磷酸基团的初始离子化和碳正离子中间体的形成。功能域中部分氨基酸一般认为是结合3个Mg2+, 触发底物上二磷酸基团离子化的关键残基,二磷酸基团离子化后, 形成活化的碳正离子中间体, 然后与双键或者水分子反应, 完成环化反应。值得注意,在所有陆生植物中参与合成植物激素的KSs中,鉴定出一个关键的氨基酸为异亮氨酸I,此氨基酸能去碳正离子中间体的质子化,对ent-karene的合成特别重要,被认为是KSs特殊PIx保守功能域(图3)。此外,根据不同物种的TPSs催化功能和重要功能域对其进行进化分析,为功能与结构的预测提供了关键的信息。

图3. KSs催化反应及关键氨基酸
虽然 TPSs 可以通过将水结合到其碳阳离子级联反应中来生成含氧产物,但它的主要功能还是合成纯烃化合物,二萜化合物的疏水性需要CYPs催化加氧以增加溶解度并提供氢键结合潜力赋予特定的生物活性。该论文通过举例论述了CYPs在不同物种中不同化学结构二萜天然产物的合成及代谢调控中起到关键作用,总结了不同物种间及同种物种间类似功能的CYPs,并阐述不同CYPs平行进化关系(图4)。
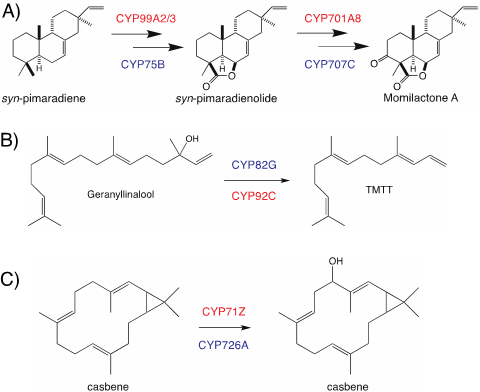
图4. 不同植物具有类似功能的CYPs
目前,对下游的修饰酶研究得还比较少。该论文以赤霉素合成最后一步关键酶:2-酮戊二酸依赖型氧化酶(2ODDs)和短链脱氢酶/还原酶(SDRs),为例阐述了下游修饰酶对二萜天然产物合成的关键作用。
最后,该论文分析发现,合成基因簇的发现和进化对二萜天然产物的合成非常有意义的。以水稻中位于4号染色体合成Momilacones基因簇、位于2号染色体合成Phytocassanes基因簇和最近发现的位于7号染色体合成基因簇为例,阐述了不同基因簇中TPSs、CYPs及下游修饰酶的功能和进化关系(图5)。此外,总结了可以合成Momilacones的不同物种(例如:水稻、稗草、小麦等),并比较分析它们之间的合成和进化差异。
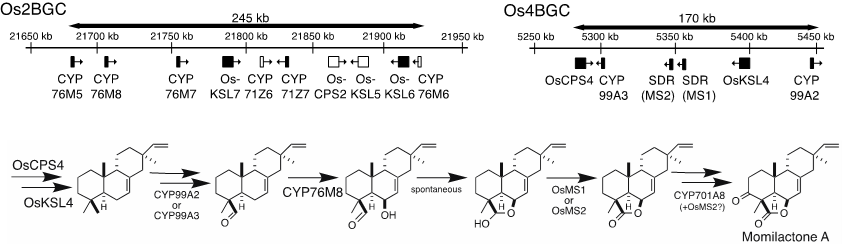
图5. 水稻Os2BGC、Os4BGC及Momilactone A合成通路
本研究主要依托美国爱荷华州立大学、北京科技大学和北京中智生物农业国际研究院等单位完成,美国爱荷华州立大学博士后及北京中医药大学助理研究员王志标为该论文第一作者,美国爱荷华州立大学Peters教授、北京科技大学万向元教授、张娟副教授为该论文共同通讯作者。同时,美国田纳西大学健康科学中心Nelson教授也参与了该工作。该研究得到美国NIH和北京科技大学青年成长国际交流计划等项目资助。
北京科技大学生物与农业研究中心、北京中智生物农业国际研究院万向元教授团队主要从事玉米等作物雄性不育机理与育种制种应用、玉米高产抗病耐逆和养分高效利用等重要性状功能基因挖掘与分子育种应用、玉米等植物营养品质及功能食品开发等方面的研究,欢迎有志优秀青年人才加入。
原文链接:https://doi.org/10.1039/D2NP00054G