


Science Bulletin | 万向元团队发现ZmSWEET6A/6B通过调控糖脂代谢平衡精准控制玉米雄花发育
糖代谢对植物雄性生殖发育至关重要,为小孢子胼胝质壁形成、初生外壁沉积、花粉内壁发育及淀粉积累等关键过程提供物质与能量支撑[1]。糖代谢相关基因发生缺陷通常会导致细胞核雄性不育(genic male sterility,GMS)。目前,植物中已鉴定出 60 多个糖代谢相关GMS 基因,主要集中在拟南芥和水稻中,而玉米中仅报道了2个[2,3],且其调控机制仍不清晰。同时,植物雄花发育过程中糖脂代谢平衡的分子机制至今还未见报道。

2026 年3月10日,北京科技大学现代农学院/生物农业研究院、北京中智生物农业国际研究院万向元团队在 Science Bulletin(IF: 21.1)在线发表题为 ZmSWEET6A/6B are required for maize male fertility via balancing sugar and lipid metabolism during pollen and anther development 的研究论文。该研究利用CRISPR/Cas9编辑技术鉴定到两个己糖转运GMS基因——ZmSWEET6A和ZmSWEET6B(均编码Sugars Will Eventually be Exported Transporters,SWEET),其双突变体表现为完全雄性不育。ZmSWEET6A/6B 均定位于细胞质膜,受转录因子ZmMYB43 直接调控,通过介导己糖转运维持花药与花粉发育中的糖脂代谢平衡,进而保证花粉正常发育,其功能丧失会导致花粉及花药发育异常。此外,还鉴定到影响ZmSWEET6A/6B转运活性的关键氨基酸位点,并证实该育性调控机制在水稻中同样保守,为作物雄性不育分子育种和制种提供了重要基因资源。
该团队利用 CRISPR/Cas9 技术创制基因编辑材料,发现ZmSWEET6A 和 ZmSWEET6B的单突变体均表现为完全可育,而双突变体(sweet6a;6b)则呈现完全雄性不育,表现为花粉外壁异常增厚、花药角质层缺失、花粉粒无法正常填充淀粉(图1a-d),表明 ZmSWEET6A 和 ZmSWEET6B在雄花发育方面存在功能冗余。进一步研究发现,二者均在花药发育的S5-S6时期的绒毡层和花粉母细胞,以及S10-S12时期的绒毡层、小孢子和发育中的花粉粒特异性表达(图1e-g);两者均定位于细胞质膜(PM)(图1h),为其发挥糖转运功能提供场所。与野生型相比,sweet6a;6b绒毡层程序性死亡(PCD)延迟至 S11 时期;S8b-S10 时期双突变体中双核绒毡层细胞比例显著降低;S11-S12 时期双突变体花药中活性氧(ROS)含量显著升高,同时伴随 24 个 ROS 清除基因显著下调、6 个 ROS 生成基因显著上调(图1i-l),表明ZmSWEET6A/6B功能缺失会导致花药中ROS稳态失衡,进而引发绒毡层降解延迟,最终导致花粉外壁异常增厚、花粉败育。
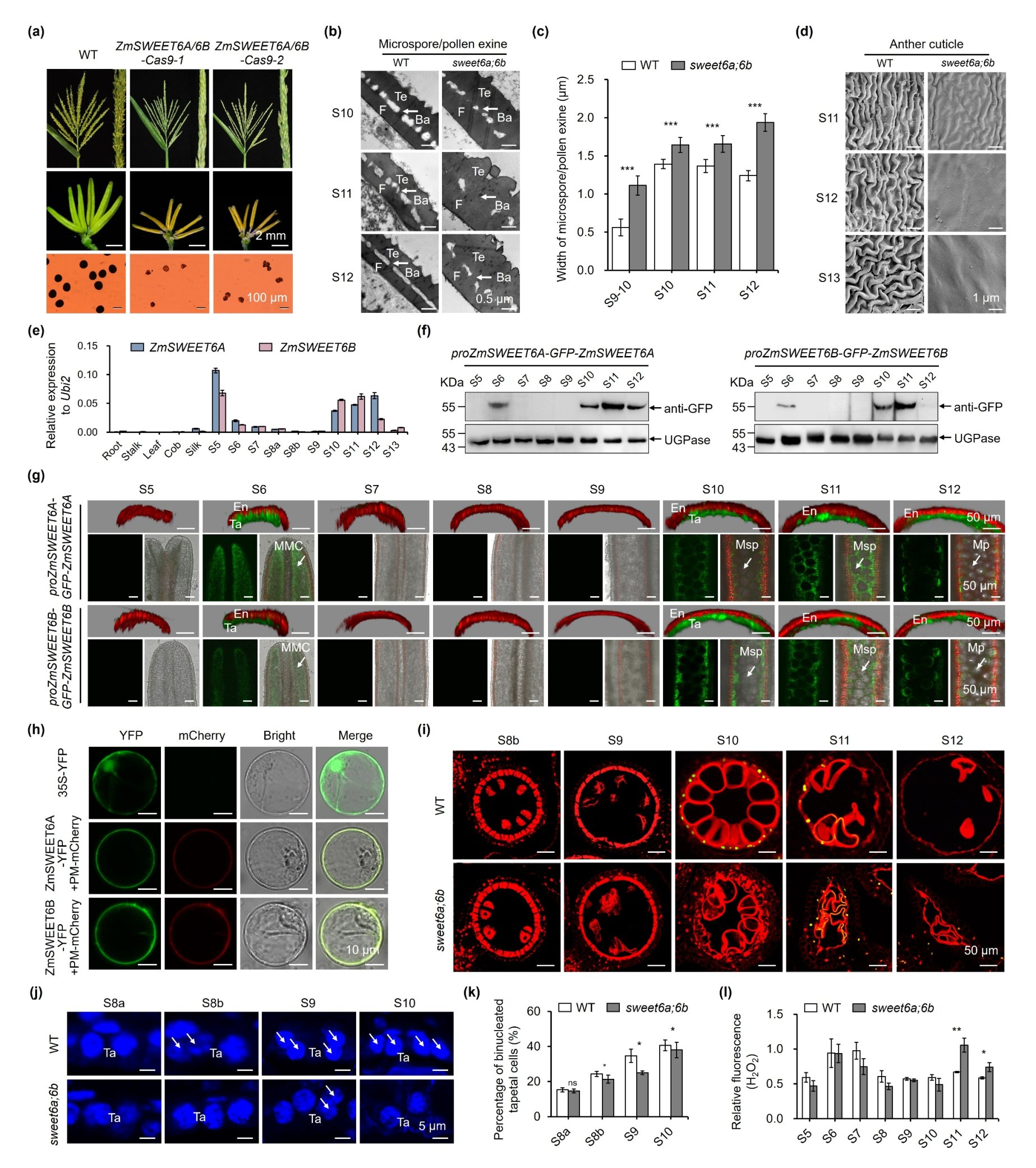
图1. sweet6a;6b双突变体绒毡层PCD延迟导致花粉外壁增厚、花药角质层光滑和雄性不育
ZmMYB43表达模式与ZmSWEET6A/6B在花药发育早期高度相似;TDLR和EMSA实验证实ZmMYB43可直接激活ZmSWEET6A/6B的转录(图2a,b)。缺陷型酵母互补和玉米原生质体糖摄取实验表明,ZmSWEET6A/6B具有己糖转运活性(主要为葡萄糖)(图2c,d),并鉴定出W57、N76、W180和N196四个对转运活性至关重要的保守残基。qRT-PCR分析显示,sweet6a;6b中ZmSWEET6A/6B在 S10–S12 时期的表达峰消失(图 2e),进一步通过PAS染色发现,sweet6a;6b花药药室内皮层(花药光合作用产生多糖的场所)中多糖颗粒显著异常积累(图2f,g),结合转录组及qRT‑PCR 分析发现,sweet6a;6b中糖代谢相关基因异常表达,其中ZmSUT5、ZmSTP8、ZmSTP11等关键糖转运基因表达下调(图 2h)。上述结果说明,ZmSWEET6A/6B功能缺失阻断了己糖从花药壁向药室外排的转运。
花粉内壁主要成分为果胶、纤维素和半纤维素 [1],而糖转运受阻会直接影响这些物质的合成。通过钙荧光白染色发现,sweet6a;6b花粉无荧光信号,表明花粉内壁完全缺失(图 2i)。综上,ZmSWEET6A/6B 介导的己糖外排是维持花药和花粉糖代谢稳态、保障花粉内壁正常形成的关键过程,该过程的紊乱会直接导致花粉发育异常。
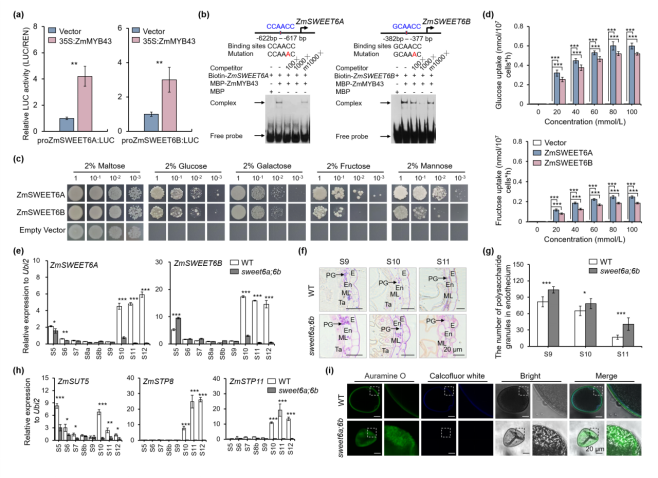
图2. ZmSWEET6A/6B 介导的己糖外排确保花粉内壁形成,对维持花药和花粉糖代谢稳态至关重要
花粉外壁和花药角质层对花药和花粉发育至关重要[4]。在sweet6a;6b中,花粉外壁(主要成分为孢粉素,S9 时期开始形成)显著增厚(图 1b,c);而花药角质层(主要成分为角质和蜡质,S10 时期开始形成)几乎完全缺失(图 1d),说明花药脂代谢异常。转录组和qRT‑PCR分析发现,sweet6a;6b中13个孢粉素合成相关基因在S5–S9 时期(尤其 S8b)表达上调;而6 个角质 / 蜡质合成相关基因在S8b–S10 时期表达下调(图 3a,b)。脂质组分析显示,sweet6a;6b花药中总角质和蜡质含量显著降低(图3c)。因此,ZmSWEET6A/6B除参与糖代谢调控外,还通过维持花药发育过程中糖与脂代谢的稳态平衡,在脂代谢中发挥关键作用;其功能缺失会同时扰乱糖、脂两大代谢通路,进而影响花药和花粉发育。
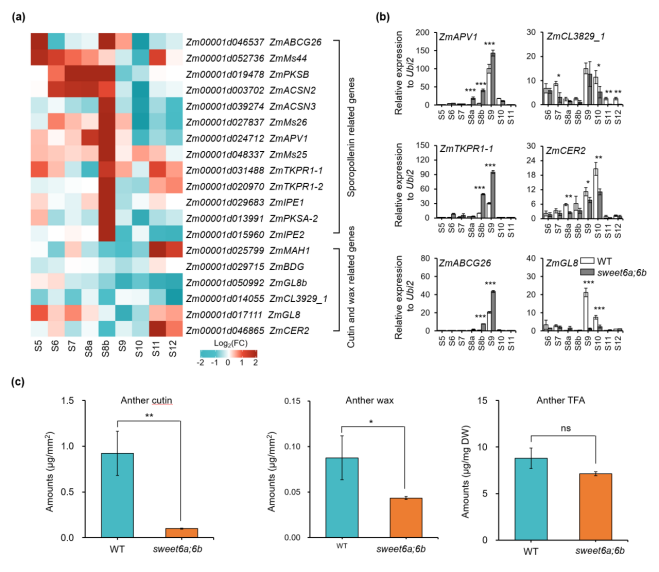
图3. ZmSWEET6A/6B对维持花药和花粉脂代谢稳态中发挥关键作用
基于上述结果,提出ZmSWEET6A/6B调控玉米雄性发育的工作模型(图 4):在花药发育过程中,ZmSWEET6A/6B 受 ZmMYB43 直接激活,成为调控代谢通路的关键因子。在野生型中,ZmSWEET6A/6B介导的己糖外排为花粉内壁形成和淀粉积累提供碳骨架,并维持 ROS 稳态;糖代谢与氧化还原状态的平衡保证了孢粉素和角质/蜡质的正常供应,进而支持花粉外壁与花药角质层正常发育。而在sweet6a;6b中,己糖外排受阻同时破坏碳源供应与 ROS 稳态,进一步干扰脂代谢平衡:孢粉素合成增多导致花粉外壁增厚,角质/蜡质合成减少导致花药角质层缺失。综上,ZmSWEET6A/6B作为代谢调控因子,通过精准调控花粉与花药发育中的糖脂代谢平衡来维持玉米正常育性。
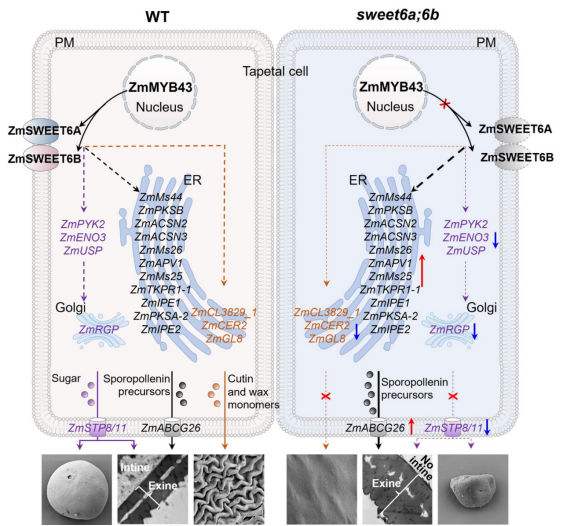
图4. ZmSWEET6A/6B通过平衡糖代谢与脂代谢保证花药和花粉正常发育的工作模型
对ZmSWEET6A/6B在单子叶和双子叶植物中进行微共线性分析发现,其同源基因在物种间高度保守。利用CRISPR/Cas9技术创制水稻OsSWEET6A/6B的单突变体和双突变体,发现仅双突变体表现为完全雄性不育(图 5),与玉米结果一致,证实SWEET6A/6B调控雄性育性的机制在水稻中保守,揭示了SWEET6类转运蛋白在作物雄性发育中的功能保守性。
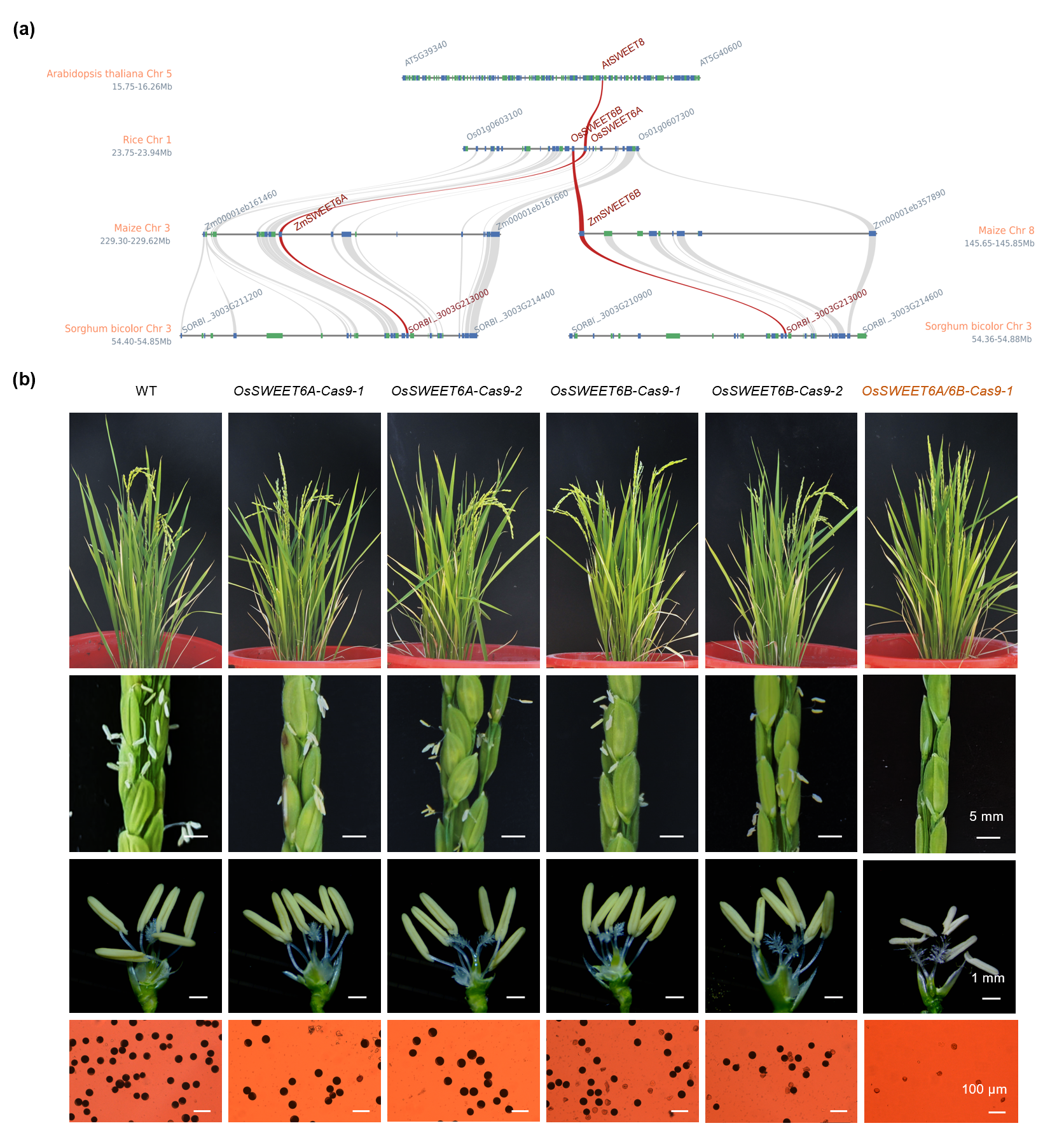
图5. SWEET6A/6B在单子叶植物水稻中的功能保守性分析
综上,ZmSWEET6A/6B作为玉米雄性发育必需的己糖转运蛋白,通过调控己糖外排维持花药物质代谢与氧化还原稳态,确保绒毡层适时发生PCD;其功能缺失会扰乱糖脂代谢稳态,最终导致玉米和水稻均表现雄性不育。该研究不仅揭示了糖脂代谢协同调控植物雄性育性的新机制,还提供了具有应用价值的玉米不育基因资源。
北京科技大学现代农学院博士生柳双双、博士后江易林为该论文共同第一作者,万向元教授和安学丽教授为该论文共同通讯作者。该研究得到了国家农业生物育种重大项目、国家重点研发计划项目、国家自然科学基金重点项目等资助。
参考文献:
[1] Liu S, Li Z, Wu S, et al. The essential roles of sugar metabolism for pollen development and male fertility in plants. Crop J 2021; 9: 1223-1236.
[2] Wang D, Skibbe D, Walbot V. Maize Male sterile 8 (Ms8), a putative β-1,3-galactosyltransferase, modulates cell division, expansion, and differentiation during early maize anther development. Plant Reprod 2013; 26: 329-338.
[3] Niu Q, Shi Z, Zhang P, et al. ZmMS39 encodes a callose synthase essential for male fertility in maize (Zea mays L.). Crop J 2023; 11: 394-404.
[4] Wan X, Wu S, Li Z, et al. Lipid Metabolism: Critical Roles in Male Fertility and Other Aspects of Reproductive Development in Plants. Mol Plant 2020; 13: 955-983.
原文链接:https://doi.org/10.1016/j.scib.2026.03.018