


PBJ | 发现 ZmMYB84-ZmPKSB调控模块控制玉米花药和花粉脂类物质分配平衡及雄配子育性
花药角质层(anther cuticle)和花粉外壁(exine)是植物雄性发育过程中形成的两个重要脂质层,它们对保护花粉正常发育和花粉-柱头互作以完成受精至关重要。花药角质层主要由角质(cutin)和蜡质(wax)组成,花粉外壁则主要由孢粉素(sporopollenin)组成,它们的主要成分都是脂质及其衍生物,均在花药绒毡层中合成,然后被运输到小孢子和花药的外表面。花药角质层和花粉外壁的脂类物质,在绒毡层中合成时,共享一些特定的脂质生物合成途径。植物需要精确控制绒毡层合成的脂质及其衍生物在花药中的正确分配,确保形成适当厚度的花药角质层和花粉外壁,抵御外界生物和非生物胁迫,最终完成双授精和产生种子。然而,这些过程中脂质分配平衡的分子机理和调控途径尚未报道。
2022年8月16日,北京科技大学、北京中智生物农业国际研究院万向元教授团队在国际期刊Plant Biotechnology Journal(IF:13.263)在线发表题为 “The ZmMYB84-ZmPKSB regulatory module controls male fertility through modulating anther cuticle — pollen exine trade-off in maize anthers”的研究论文,在玉米中首次发现ZmMYB84-ZmPKSB调控模块控制玉米花药角质层和花粉外壁脂类物质分配平衡,并决定雄配子(花粉)发育和育性,加深了对植物精准调控花药角质层和花粉外壁脂质分配过程的认识。

该团队发现了一个特异的玉米雄性不育突变体pksb,表现为花粉外壁变薄,但花药角质层增厚和变密(图1)。通过图位克隆的方法分离到该突变体的育性恢复基因ZmPKSB,利用CRISPR/Cas9介导的基因编辑方法对目标候选基因进行了功能验证。表达分析发现ZmPKSB在S8b至S9-10时期的玉米花药中特异表达,在S9时期表达最高;ZmPKSB基因编码一个内质网定位的聚酮合酶(polyketide synthase:PKS)。进一步酶活性分析发现,ZmPKSB重组蛋白可以催化丙二酰辅酶A与中链脂肪酰辅酶A,生成三酮和四酮α-吡喃酮(triketide和tetraketide α-pyrone),并且其酶活性功能依赖于一个保守的催化三联体(C171、H320和N353)(图2)。

图1. 玉米pksb突变体与野生型(WT)的表型和细胞学比较
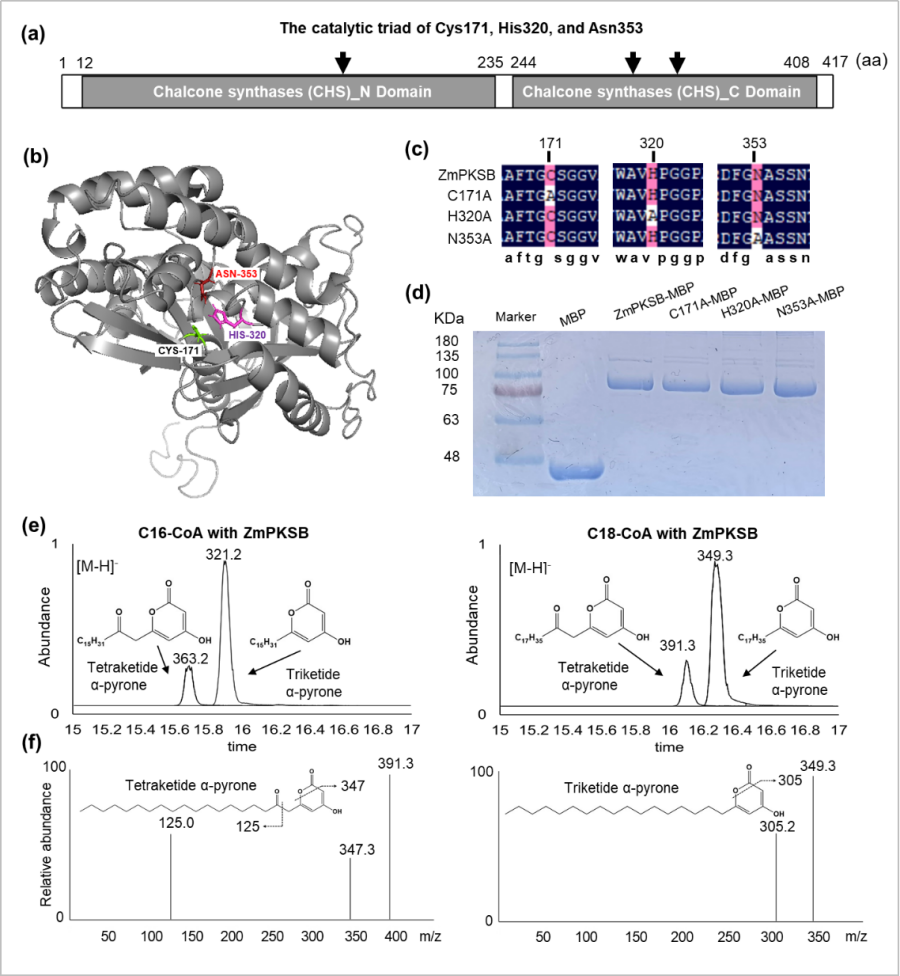
图2. ZmPKSB酶活性依赖一个保守的催化三联体(C171、H320和N353)
然后,qPCR和生化实验表明ZmPKSB表达由转录因子ZmMYB84直接激活,并且ZmMYB84功能丧失后也导致花药角质层更致密但花粉外壁更薄,与突变体pksb的表型相似(图3)。pksb和myb84突变体花粉外壁变薄但花药外壁变密,暗示着孢粉素的生物合成和运输可能受到抑制,但角质和蜡质的生物合成和运输过程可能被增强。进一步分析9个孢粉素相关基因与9个角质和蜡质相关基因,分别在pksb和myb84突变体不同发育时期花药中的表达模式,结果发现:与野生相比,pksb突变体中所有孢粉素合成相关基因的表达在S9时期均显著下调,而所有角质和蜡质相关基因的表达在S10时期则显著上调;类似结果也在myb84突变体中被检测到。此外,pksb和myb84突变体花药中角质和蜡质的总含量和主要单体含量也都类似地较野生型显著增加。因此,在细胞学、基因转录和脂质代谢三个水平上,ZmMYB84和ZmPKSB完全同步控制花药角质层和花粉外壁形成,二者形成一个控制脂类物质分配的调控模块(图3h)。
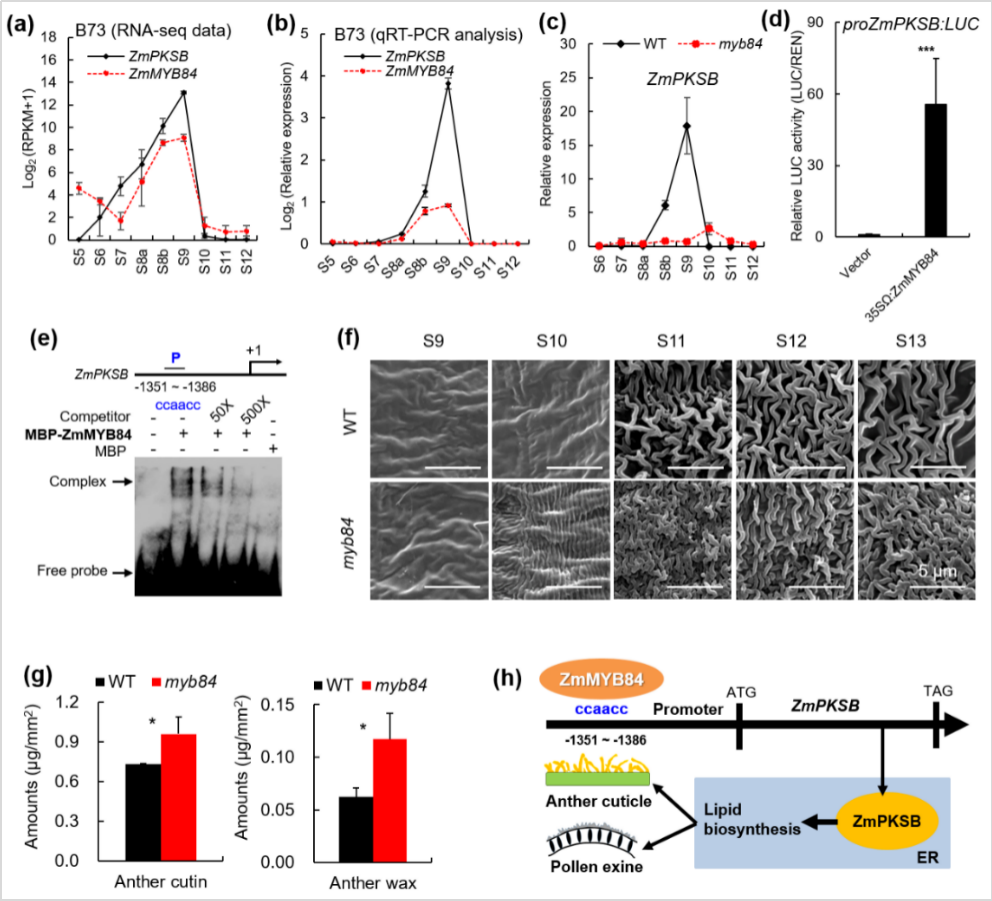
图3. ZmMYB84-ZmPKSB调控模块控制花药角质层和花粉外壁脂质生物合成
基于上述结果,研究团队提出了ZmMYB84-ZmPKSB调控模块的工作模型:ZmMYB84-ZmPKSB模块通过同步调控花药绒毡层中与孢粉素、角质和蜡质的生物合成和运输相关的一系列基因,进而控制花药角质层和花粉外壁形成所需脂类物质的分配平衡(图4)。该工作模型的建立为“植物精准调节脂类物质代谢平衡,确保形成适当厚度的花药角质层和花粉外壁,抵御外界生物和非生物胁迫,最终完成双授精和产生种子”提供了新见解。
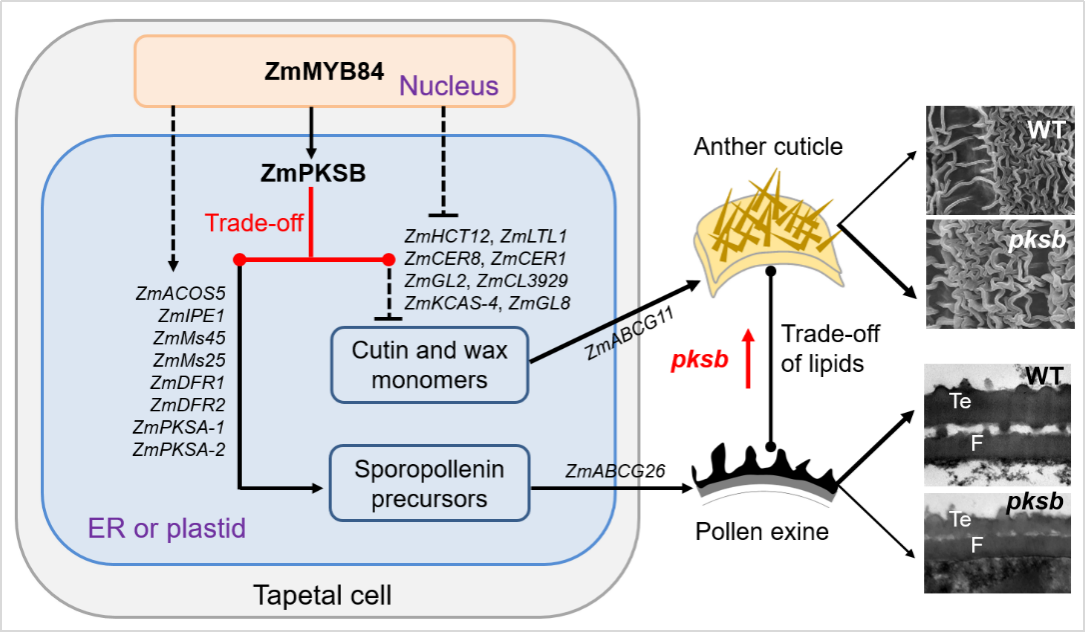
图4. ZmMYB84-ZmPKSB调控模块控制“玉米花药角质层和花粉外壁脂类物质分配平衡及雄配子育性”的工作模型
本研究主要依托北京科技大学生物与农业研究中心和北京中智生物农业国际研究院等单位完成,北京科技大学博士生刘欣泽和博士生江易林为该论文共同第一作者,万向元教授和安学丽教授为共同通讯作者。该研究得到国家重点研发计划、国家自然科学基金、中央高校基本科研业务费、北京市科技计划等项目联合资助。
原文链接:https://doi.org/10.1111/pbi.13911